Unsere diesjährige YAQU PACHA Jahreshauptversammlung fand am Freitag, den 26.05.2023 von 18:30 bis 20:30 Uhr im Naturkundehaus des Tiergarten Nürnberg statt und auch via Zoom Konferenz nahmen unsere Biologen und Mitglieder der Gesellschaft an der Veranstaltung teil.

Dr. Lorenzo von Fersen (1. Vorsitzender YAQU PACHA e.V.) führte durch die Veranstaltung. Nach einer kurzen Einleitung berichteten zunächst unsere Biologen aus Lateinamerika über die Arbeit an ihren Projekten.

Prof. Eduardo Secchi begann mit der Vorstellung der Projekte, die mit unseren Partnern von ECOMEGA FURG von der Bundesuniversität von Rio Grande FURG in Brasilien durchgeführt werden. Besonderes Augenmerk liegt hier auf der Zusammenarbeit mit Fischergemeinden und Schulen, um langfristig den Beifang von Meeressäugern zu reduzieren und den Kontakt mit den Fischern zu intensivieren. Ziel des Projektes ist es ebenfalls, das Thema Natur- und Artenschutz langfristig in den Unterricht in Schulen zu integrieren. Das Team um Prof. Secchi arbeitet auch am Aufbau eines Hormonphysiologischen Labors für die Analyse von Cortisol und anderen Hormonen von Proben der dort lebenden Tierarten. Dr. Silvina Botta nahm an einer Schulung im weltweit renommierten Labor des Toronto Zoo von Dr. Mastromonaco teil.

Dr. Lorenzo Rojas-Bracho berichtete im Anschluss über die Arbeit zum Schutz des vom Aussterben bedrohten Vaquita. Er stellte die Arbeit von PESCA ABC vor, die alternative Fischereimethoden durchführen, um damit den Beifang von Vaquitas zu verhindern. Durch die Regierung Mexikos wurden im Verbreitungsgebiet der Vaquitas ebenfalls schwere Betonblöcke mit Metallhaken versenkt, um die Fischerei in diesem Gebiet zu verhindern, da die Netze durch diese Metallhaken zerstört werden würden. Offensichtlich hat dies bereits einen positiven Effekt und die Fischer scheinen diese Gebiete zu meiden. In den letzten 3 Wochen hat das Team um Dr. Rojas-Bracho wieder Expeditionen unternommen, um Vaquitas zu zählen und es gab wieder Sichtungen von Tieren, was uns positiv stimmt.
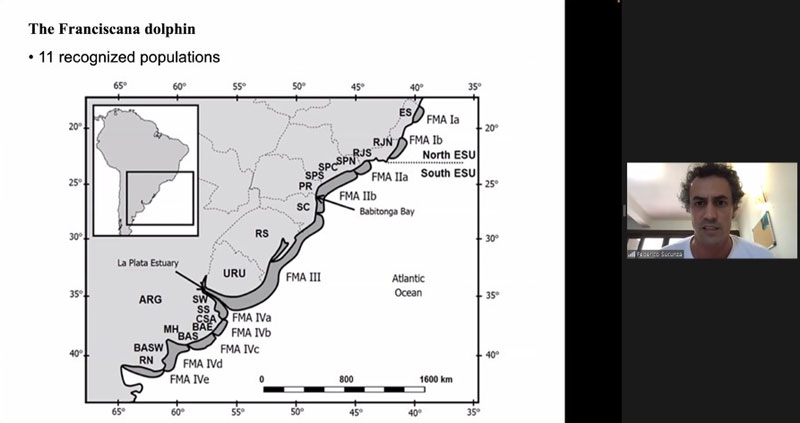
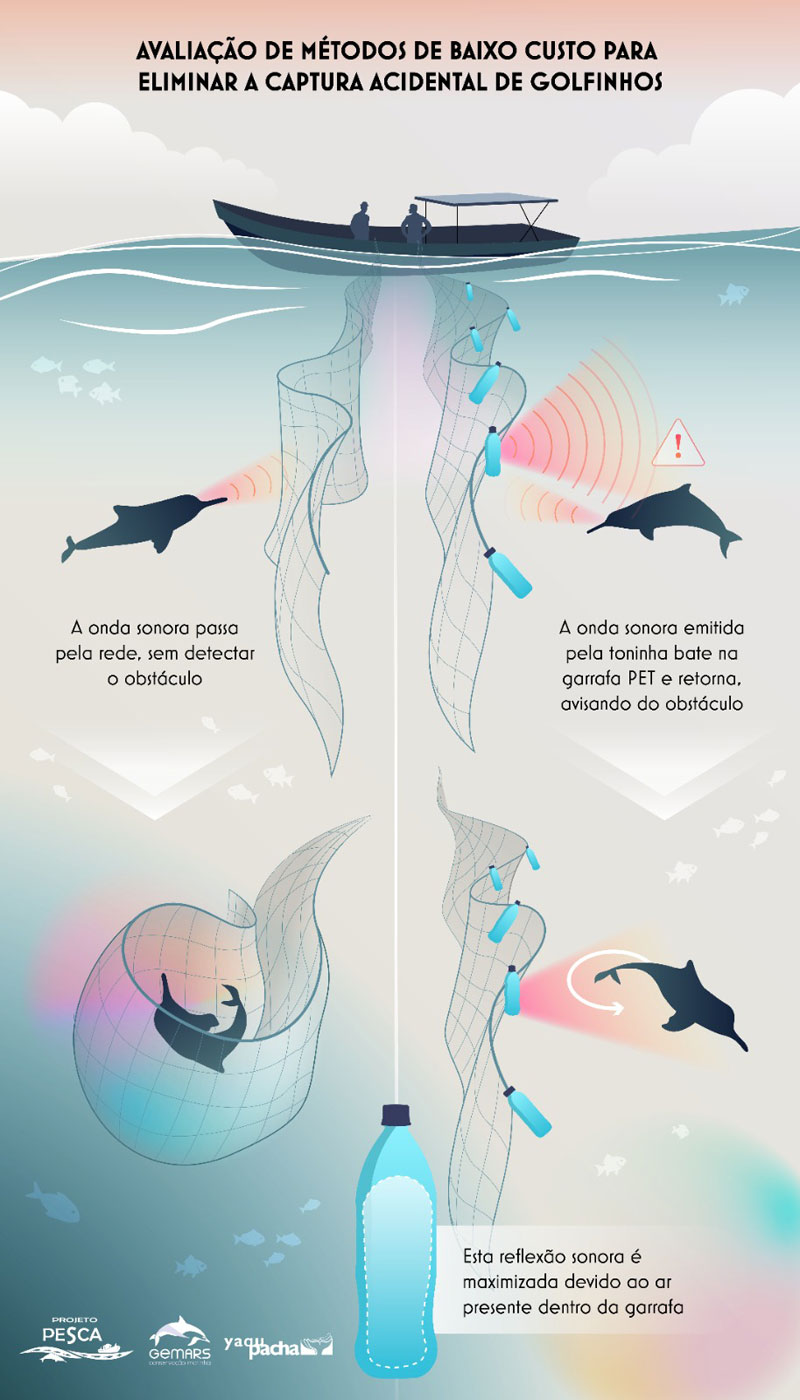
Dann stellte Dr. Federico Sucunza von unseren Partnern GEMARS und INSTITUTO AQUALIE seine Arbeit beim Schutz des La Plata Delfins – Franciscana – Toninha vor und berichtete über die Flugzählungen, die im März und April zum ersten Mal in Uruguay durchgeführt wurden. Nach Auswertung der Daten, kann man derzeit von einem Bestand von etwa 30000 Franciscanas in Uruguay ausgehen, wobei es dort eine sehr hohe Sterblichkeit durch Beifang gibt. Um dem entgegenzuwirken, arbeiten wir zusammen an der Anbringung von akustischen Signalen, wie Pinger. Dr. Federico Sucunza berichtete auch über unsere Zusammenarbeit mit dem Team von PROJETO PESCA, um eine einfache und kostengünstige Alternative zu den Pingern zu testen und zu etablieren. Es zeigt sich, dass leere Plastikflaschen, die in den Netzen montiert werden, von den Delfinen wahrgenommen werden, sodass diese die Netze meiden. Erste Tests verliefen sehr erfolgreich.

Im Anschluss berichtete unsere Biologin Yurasi Briceño von unseren Partnern PROYECTO SOTALIA über ihre Arbeit beim Schutz der Flussdelfine Inia geoffensis und Sotalia und den Manatis in Venezuela. Die Erdölförderung im Maracaibo See und die Gewinnung von Gold mit Hilfe von Quecksilber stellen eine ernsthafte Bedrohung für die Tiere dar und es zeigt sich, dass sich das Quecksilber in den Fischen so stark anreichert, sodass es auch über den Nahrungskreislauf für die Menschen zu einer ernsthaften Gefahr wird. Die Arbeit des Teams um Yurasi Briceño erstreckt sich über viele Bereiche, wie zum Beispiel regelmäßige Bestandszählungen der Tiere und ein wichtiger Bereich bildet auch die Umweltpädagogik.

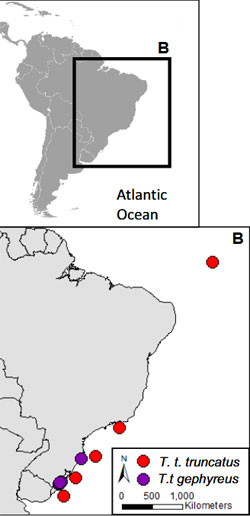
Nachfolgend berichtete Dr. von Fersen über die Maßnahmen, die speziell zum Schutz des bedrohten Lahille’s Großen Tümmlers (Tursiops truncatus gephyreus) in Brasilien durchgeführt werden.
Ein weiterer Schwerpunkt der Arbeit von YAQU PACHA ist das sogenannte Capacity Building, der Aufbau von Kompetenzen und die Schulung und Weiterbildung unserer Mitarbeiter.
YAQU PACHA war auch entscheidend beim Aufbau der Alliance for Franciscana Dolphin beteiligt, die in verschiedenen Arbeitsgruppen Schutzkonzepte für den bedrohten La Plata Delfin – Franciscana – Toninha (Pontoporia blainvillei) entwickelt.
Das Latin American Journal for Aquatic Mammals LAJAM blickte auf sein 20-jähriges Bestehen zurück und YAQU PACHA ist seit Beginn an Hauptsponsor dieses wichtigen Wissenschaftsmagazins.
Im Jahr 2022 war YAQU PACHA ebenfalls Hauptsponsor der 19. RT der SOLAMAC, dem wichtigsten Arbeitstreffen für Biologen, die am Schutz von wasserlebenden Säugetierarten in Lateinamerika arbeiten.
Herr Dr. von Fersen berichtete auch über den großen internationalen Workshop Human Dimension in small Cetacean Conservation, der von YAQU PACHA organisiert und durchgeführt wurde. Ein finaler Abschlussbericht folgt in kürze.
Ein weiterer wichtiger Teil unserer Arbeit war die Zusammenarbeit mit der Gruppe ECOLOGIA HUMANA DO OCEANO, die von unserer Biologin Camilah Antunes Zappes in Brasilien geleitet wird und deren Schwerpunkt der Zusammenhang von Mensch und Umwelt ist. Hauptaufgabengebiet der Gruppe ist die Umweltbildung in Schulen und Kindergärten und die Kommunikation mit Fischergemeinden.
Sandra Honigs (2. Vorsitzende, YAQU PACHA e.V.) stellte die Arbeit und die Aktivitäten von YAQU PACHA in Deutschland, bzw. Europa vor. An dieser Stelle ein herzliches Dankeschön an alle ehrenamtlichen Aktiven, ohne deren Hilfe unsere Veranstaltungen und Aktionen nicht möglich wären.
Zum Abschluss stellte Dr. Lorenzo von Fersen noch das neue Programm YAQU PACHA for Diversity vor, das am Beispiel des Lahille’s Großen Tümmlers (derzeitiger Bestand 600 Tiere) zeigt, weshalb es so wichtig ist, die biologische Vielfalt zu erhalten und zu schützen.
Das ausführliche Protokoll zur Jahreshauptversammlung mit allen Aktivitäten unserer Organisation finden Sie in kürze ebenfalls hier auf unserer Website.